磁性細菌 (Magnetotactic bacteria)
磁性細菌研究の背景
地磁気を感知する細菌
磁性細菌は、外界から鉄イオンを細胞の中に取りこみ、マグネタイト(磁鉄鉱)とよばれる鉱物の結晶を作ります。そして、この磁鉄鉱の結晶を直鎖状に並べ、棒磁石のようにして細胞の中心に配置します。この細胞内構造は、マグネトソームとよばれており、磁性細菌はマグネトソームをコンパスのように用いて、磁気(地磁気)を感知することができます。
磁鉄鉱の結晶を被う膜は、細胞質膜の貫入により形成されます。また、細胞骨格の繊維がマグネトソーム鎖に沿って配置されています。このように、マグネトソームはとても複雑な構造をしています。マグネトソームは、植物細胞や動物細胞がもつオルガネラと同じように、生体膜で区切られ、特異的な機能をもち、細胞分裂を介して娘細胞へ伝達されます。このことから、マグネトソームは、原核細胞のオルガネラとよばれています。私たちはマグネトソームの構造とその機能を明らかにし、原核細胞の細胞骨格やオルガネラの働きや形成過程や、生物鉱物をつくる過程(バイオミネラリゼーション)、生物の磁気感知の仕組みの解明をめざしています。
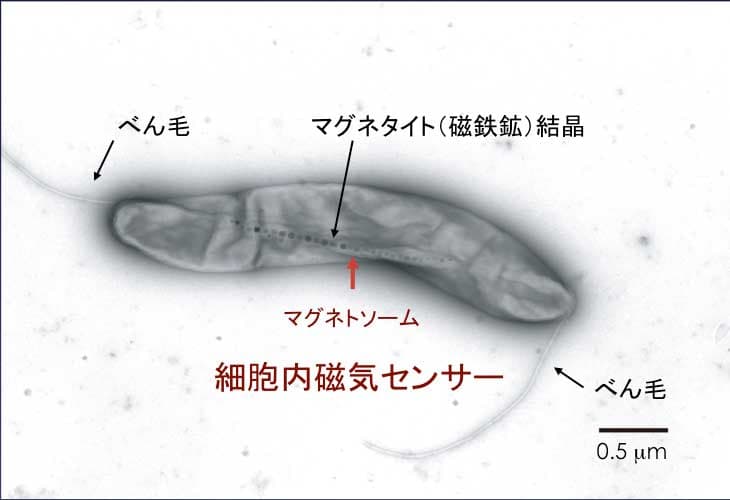
磁性細菌のオルガネラ「マグネトソーム」の研究
マグネトソームは、細胞質膜が陥入することで生じたリン脂質の膜小胞内に磁性鉱物の結晶を合成することで形成されます。マグネトソームは、細胞骨格のはたらきにより細胞中央に直鎖状に配置され、棒磁石のような構造となり効率的な磁気センサーとして機能します。磁性細菌の形成する磁気微粒子は、均質な結晶型をしており、単磁区構造を保持する大きさ(約35~100 nm)に制御されており、効率的な磁石として機能します。マグネトソームには、細胞内の他の部位には存在しない数十種類の蛋白質が特異的に局在しており、マグネトソームの形成や機能を担っていますが、そのメカニズムはほとんど分かっていません。私たちは、微生物学、生化学、分子生物学的な手法と、高速原子間力顕微鏡(高速AFM)、生細胞蛍光イメージング、電子顕微鏡技術といった最新のイメージング技術を組み合わせ、マグネトソームの構造やマグネトソーム蛋白質の機能に関する研究を行っています。




当研究室における磁性細菌の研究
マグネトソームの構造とMamA
マグネトソームの形成や維持に関わる蛋白質の遺伝子は、磁性細菌のゲノムの特定の領域 ‘Magnetosome Island (MAI)’ にまとまってコードされている。MAIに存在するもっとも大きいmamAB オペロンにはMamH、 I、 E、 J、 K、 L、 M、 N、 O、 P、 A、 Q、 R、 B、 S、 T、 U、 V の18種類の蛋白質がコードされ、類似した構造のオペロンがすべての磁性細菌で保存されていることから、マグネトソーム形成に必須なオペロンと考えられている。なかでも、MamAは塩基配列が決定されたすべての磁性細菌のMAIに保存されており、発現量も多い事から重要な役割を担っていると予想されていた。
当研究室のOkudaらは1996年に世界で初めてマグネトソーム蛋白質の遺伝子を決定し、MamA(当時はMam22とよばれた)がTPR (TetratricoPeptide Repeat)モチーフ(蛋白質間相互作用を担う構造モチーフ)をもつ蛋白質であることを発表した(Gene (1996) 171:99-102, FEBS Lett. (2001) 491:169-173)。また、私たちは、MamAの局在を免疫電子顕微鏡法により検討した。その結果、MamAはマグネトソーム膜の外側(細胞質側)に局在することを明らかとした(J. Bacteriol. (2006) 188:3805-3812)。さらに、2010年には、高速AFMを用いてマグネトソームを観察することに成功した(PNAS (2010) 107:9382-9387)。高速AFMは、生理的条件下で機能中の生体分子の構造とダイナミクスを、高い空間時間分解能で記録できる走査プローブ顕微鏡の一種であり、水溶液中で試料表面を先端を尖らせた針で走査することにより表面形状を原子レベルの分解能で観察することができる(詳細は金沢大学理工研究域バイオAFM先端研究センターホームページを参照)。AFMによる解析の結果、マグネトソームは、約7 nmの厚さの有機層で被われていることを明らかにした。また、MamAはマグネタイトを覆うリン脂質二重膜の外側で蛋白質の殻を形成し、マグネトソームを被う有機層を形成している事が示唆された。
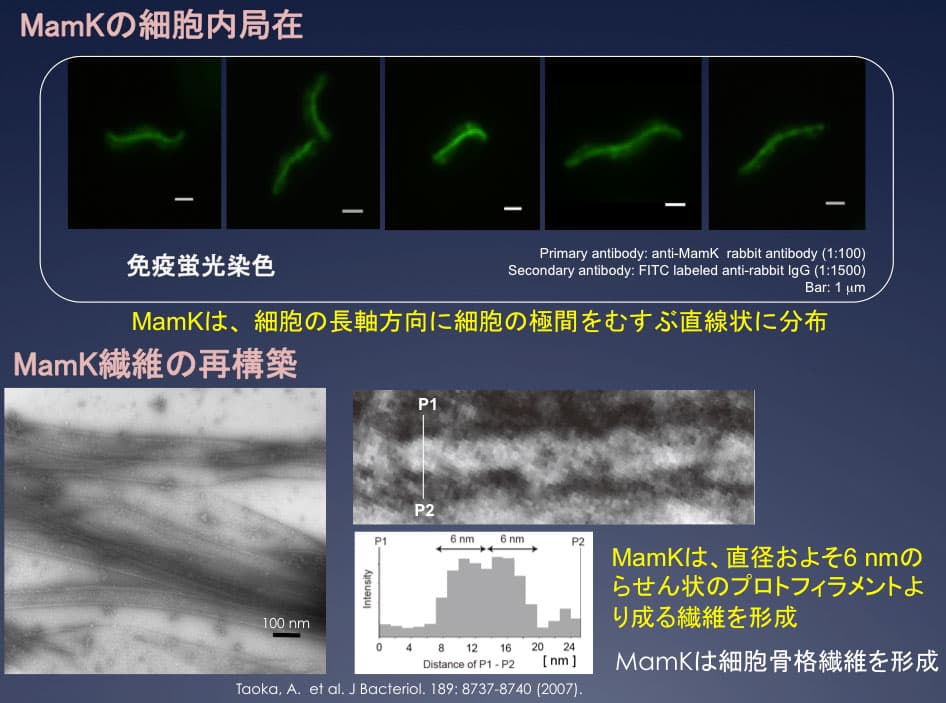
磁性細菌の細胞骨格MamK
mamABオペロンには、アクチンホモログであるMamK蛋白質がコードされている。MamKの細胞内局在を調べたところ、MamK は細胞両極にまで達する細胞長軸に沿った直線的な繊維状の構造を形成していることを確認した。大腸菌で発現、精製した単量体のMamK蛋白質は、直線的な束状のMamK細胞骨格繊維に重合することを確認した。MamK繊維は、プロトフィラメントとみられる直径 6 nmほどのらせん状の細いフィラメントが絡まり合って構成されていることを報告した(J Bacteriol. (2007) 89:8737-8740)。
従来の走磁性測定法である光散乱法ではmamK欠損株と野生株との間に表現型の差異は検出されない。そこで、私たちはMamK細胞骨格の生理的機能を明らかにするため細胞の磁性と運動性を同時に評価する新しい走磁性の評価法magnetic swimming assayを開発し、MamK細胞骨格の機能解析を行った。その結果、mamK欠損株ではマグネトソームの娘細胞への安定的な分配が行われていないことを示唆され、MamK細胞骨格の新しい機能が示唆された。また、本法はMamK以外のマグネトソーム蛋白質の機能解析への利用も期待できる。(Biosci. Biotech. Biochem. (2013) 77:940-947)
マグネタイト合成に関わる新規シトクロムMamP
マグネタイトは、酸素とFe2+、Fe3+の化合物であり、細胞内におけるその生成には、鉄の酸化還元反応が必要と考えられる。MAIには、MamPとよばれるヘムc結合モチーフをもつ膜蛋白質が保存されている。そこで、MamPの機能解析を行ったところ、MamPは対数増殖期にマグネトソーム内の磁鉄鉱結晶の成長に必須であること、磁鉄鉱の形成にはMamPへのヘムcの結合が必要であることが示され、MamPは磁性鉱物の合成に関わる新奇のヘム蛋白質であることが明らかになった(FEMS Microbiol. Lett. (2014) 358:21-29)。
新規走磁性細菌の単離・培養
本研究室では磁性細菌の多様性を明らかにし、走磁性の生理的意義を解明するため新規の磁性細菌の単離培養を進めている。金沢市内の湖沼から、長さ約13 µm、直径約8 µmの巨大な磁性桿菌GRS-1株(既知の磁性細菌で最大)を見いだした。本細菌は、γプロテオバクテリアに属し、数百個以上の磁鉄鉱結晶からなる長いマグネトソーム鎖を有し、また、細胞内にカルシウムを多量に蓄えていた。細胞極で数本のべん毛を束ね、これを用いて走磁性運動を行っている事がわかった(Microbiology (2014) 160:2226-2234)。観察が容易かつ大量の磁気微粒子を合成するGRS-1を単離培養できれば、走磁性運動や磁気感応マシナリーの解明への貢献が期待される。
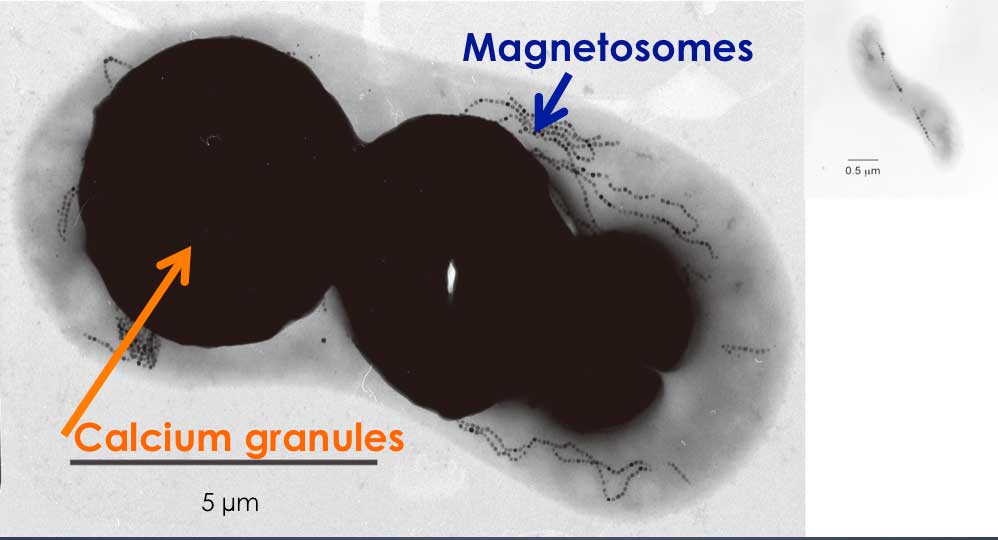
同じ縮尺の一般的な磁性細菌の透過型電子顕微鏡写真。
GRS-1は細胞内に多数のマグネトソームとカルシウムを含む巨大な顆粒をもつ。
磁性細菌のエネルギー代謝
磁性細菌のエネルギー獲得機構を明らかにするため、磁性細菌M. magnetotacticumよりチトクロムc550(Arch Microbiol. (1995) 163: 400-406),cbb3型チトクロム酸化酵素(FEBS Lett. (1994) 347: 22-26),亜硝酸塩還元酵素(cytochrome cd1) (Eur J Biochem. (1995) 233: 665-671),硝酸塩還元酵素(Canadian Journal of Microbiology (2003) 49: 197-206, 新規ヘム蛋白質(cytochrome a1 like hemoprotein) (J. Inorg. Biochem. (2000) 82: 73-78., Biochim Biophys Acta. (1993) 1158:237-243.)を精製し、その特徴を調べた。その結果、磁性細菌は微好気的酸素呼吸を行い生育に必要なエネルギーを獲得する一方で、嫌気呼吸経路である脱窒経路の酵素群も同時に発現していることが明らかになった。脱窒経路を担う酵素である亜硝酸塩還元酵素(cytochrome cd1)が新規のFe(II)酸化活性を持つことや、硝酸塩還元酵素活性が最適なマグネタイト合成に必要であることを明らかにし、脱窒経路とマグネタイト合成の関連を示唆した。また、私たちは磁性細菌がもつ新規のヘム蛋白質を発見しており、その機能が注目される。
磁性細菌に関する研究成果
- Measuring magnetosomal pH of the magnetotactic bacterium Magnetospirillum magneticum AMB-1 using pH-sensitive fluorescent proteins.
- Eguchi Y, Fukumori Y, Taoka A.
- Biosci Biotechnol Biochem. (2018) DOI: 10.1080/09168451.2018.1451739.
- 細菌の磁気感応運動のためのオルガネラ「マグネトソーム」
- 田岡東、福森義宏
- 生物工学 (2018) 96(5):248-252.
- Tethered Magnets Are the Key to Magnetotaxis: Direct Observations of Magnetospirillum magneticum AMB-1 Show that MamK Distributes Magnetosome Organelles Equally to Daughter Cells.
- Taoka A, Kiyokawa A, Uesugi C, Kikuchi Y, Oestreicher Z, Morii K, Eguchi Y, Fukumori Y.
- mBio. (2017) 8(4). pii: e00679-17. DOI: 10.1128/mBio.00679-17.
- A protein-protein interaction in magnetosomes: TPR protein MamA interacts with an Mms6 protein.
- Nguyen HV, Suzuki E, Oestreicher Z, Minamide H, Endoh H, Fukumori Y, Taoka A.
- Biochem Biophys Rep. (2016) DOI: 10.1016/j.bbrep.2016.05.010.
- 高速原子間力顕微鏡を用いたバクテリアの生細胞イメージング
- 田岡 東、福森義宏
- 化学と生物(農芸化学会誌)(2015) 53(5):293-298.
- A magnetosome-associated cytochrome MamP is critical for magnetite crystal growth during the exponential growth phase.
- Taoka A, Eguchi Y, Mise S, Oestreicher Z, Uno F, Fukumori Y.
- FEMS Microbiol Lett. (2014) 358:21-29. DOI: 10.1111/1574-6968.12541.
- Characterization of uncultured giant rod-shaped magnetotactic Gammaproteobacteria from a freshwater pond in Kanazawa, Japan.
- Taoka A, Kondo J, Oestreicher Z, Fukumori Y.
- Microbiology-SGM. (2014) 160:2226-2234. DOI: 10.1099/mic.0.078717-0.
- 磁性細菌オルガネラ「マグネトソーム」の構造機能相関の解明
- 福森 義宏、田岡 東
- 生物物理. (2014) 54(1):11-14.
- Analysis of magnetotactic behavior by swimming assay.
- Sakaguchi S, Taoka A, Fukumori Y.
- Biosci Biotechnol Biochem. (2013) 77(5):940-947. DOI: 10.1271/bbb.120880.
- Single-molecule imaging on living bacterial cell surface by high-speed AFM.
- Yamashita H, Taoka A, Uchihashi T, Asano T, Ando T, Fukumori Y.
- J Mol Biol. (2012) 422(2):300-309. DOI: 10.1016/j.jmb.2012.05.018.
- Visualization and structural analysis of the bacterial magnetic organelle magnetosome using atomic force microscopy.
- Yamamoto, D., Taoka, A., Uchihashi, T., Sasaki, H., Watanabe, H., Ando, T., and Fukumori, Y.
- Proc Natl Acad Sci U S A. (2010) 107(20), 9382-9387. DOI: 10.1073/pnas.1001870107.
- Identification of iron transporters expressed in the magnetotactic bacterium Magnetospirillum magnetotacticum.
- Taoka A, Umeyama C, and Fukumori Y.
- Curr. Microbiol. (2009) 58: 177-181. DOI: 10.1007/s00284-008-9305-7.
- 特集:原核細胞の細胞骨格「特集に当たって」
- 福森義宏.
- 蛋白質 核酸 酵素 (2008) vol.53 No.13,p1719 - p1722.
- 磁性細菌のオルガネラ、マグネトソームの細胞内局在と細胞骨格
- 福森義宏、 田岡東.
- 蛋白質 核酸 酵素 (2008) vol.53 No.13,p1746 - p1751.
- 磁性細菌のマグネタイト微粒子の形成機構
- 福森義宏、 田岡東.
- 月刊「バイオインダストリー」 (2007) vol.24(12), p5 - p13
- Polymerization of the Actin-Like Protein MamK, Which Is Associated with Magnetosomes.
- Taoka A, Asada R, Wu L-F, Fukumori Y.
- J Bacteriol. (2007) 189(23): 8737-8740. DOI: 10.1128/JB.00899-07.
- 細菌の磁気オルガネラの構造と細胞内局在
- 福森義宏、 田岡東.
- 化学と生物 (2007) 45(3):154-156.
- Biogenesis of actin-like bacterial cytoskeletal filaments destined for positioning prokaryotic magnetic organelles.
- Pradel N, Santini CL, Bernadac A, Fukumori Y, Wu LF.
- Proc Natl Acad Sci U S A. (2006) 103(46):17485-17489. DOI: 10.1073/pnas.0603760103.
- Spatial localizations of Mam22 and Mam12 in the magnetosomes of Magnetospirillum magnetotacticum.
- Taoka, A., Asada, R., Sasaki, H., Anzawa, K., Wu, L.-F., Fukumori, Y.
- J Bacteriol. (2006) 188(11):3805-3812. DOI: 10.1128/JB.00020-06.
- Enzymes for magnetite synthesis in Magnetospirillum magnetotacticum
- Fukumori, Y.
- “Biomineralization” (E.Baeuerlein ed.) WILEY-VCH. p75-90.
- Nitrate reductase from the magnetotactic bacterium Magnetospirillum magnetotacticum MS-1:purification and sequence analyses
- Taoka, A., Yoshimatsu, K., Kanemori, M., Fukumori, Y.
- Can. J. Microbiol. (2003) 49:197-206. DOI: 10.1139/w03-028.
- Expression and characterization of a magntosome-associated protein, TPR-containing MAM22, in Escherichia coli
- Y.Okuda and Y.Fukumori.
- FEBS Letters. (2001) 491:169-173. DOI: 10.1016/S0014-5793(01)02178-0.
- Heme-copper oxidase family structure of Magnetospirillum magnetotacticum 'cytochrome a1'-like hemoprotein without cytochrome c oxidase activity.
- Tanimura Y, Fukumori Y.
- J Inorg Biochem. (2000) 82:73-78. DOI: 10.1016/S0162-0134(00)00152-5.
- ナノサイズ生物磁気微粒子の構造と機能
- 福森 義宏
- 生化学 (2000) 72:1165-1172.
- Iron reductase for magnetite synthesis in the magnetotactic bacterium Magnetospirillum magnetotacticum
- Noguchi Y, Fujiwara T, Yoshimatsu K, Fukumori Y.
- J Bacteriol. (1999) 181: 2142-2147.
- Cloning and sequencing of a gene encoding a new member of the tetratricopeptide protein family from magnetosomes of Magnetospirillum magnetotacticum.
- Okuda Y, K Denda, Fukumori Y.
- Gene. (1996) 171: 99-102. DOI: 10.1016/0378-1119(95)00008-9.
- Nitrite reductase from the magnetotactic bacterium Magnetospirillum magnetotacticum. A novel cytochrome cd1 with Fe(II):nitrite oxidoreductase activity.
- Yamazaki T, Oyanagi H, Fujiwara T, Fukumori Y.
- Eur J Biochem. (1995) 233: 665-671. DOI: 10.1111/j.1432-1033.1995.665_2.x.
- Purification, primary structure, and evolution of cytochrome c-550 from the magnetic bacterium, Magnetospirillum magnetotacticum.
- Yoshimatsu K, Fujiwara T, Fukumori Y.
- Arch Microbiol. (1995) 163:400-406. DOI: 10.1007/BF00272128.
- 磁性細菌の生化学
- 福森 義宏.
- 蛋白質・核酸・酵素 (1995) 40:919-925.
- Purification, and some molecular and enzymatic features of a novel ccb-type cytochrome c oxidase from a microaerobic denitrifier, Magnetospirillum magnetotacticum.
- Tamegai H, Fukumori Y.
- FEBS Lett. (1994) 347: 22-26. DOI: 10.1016/0014-5793(94)00500-1.
- Purification and properties of a 'cytochrome a1'-like hemoprotein from a magnetotactic bacterium, Aquaspirillum magnetotacticum.
- Tamegai H, Yamanaka T, Fukumori Y.
- Biochim Biophys Acta. (1993) 1158:237-243. DOI: 10.1016/0304-4165(93)90020-9.
- Oxygen concentration-dependent induction of a 140-kDa protein in magnetic bacterium Magnetospirillum magnetotacticum MS-1.
- Sakaguchi H, Hagiwara H, Fukumori Y, Tamaura Y, Funaki M, Hirose S.
- FEMS Microbiol Lett. (1993) 107:169-174. DOI: 10.1111/j.1574-6968.1993.tb06025.x.