

有鬚動物とは、口・肛門・消化管が無く細長い触手を頭部に持つ左右対称の真体腔動物のことであり、環形動物の仲間である。例えば、ガラパゴス沖の深海(水深約1500〜2500m)の熱水噴出口付近に棲息するRiftia pachyptila(図1:ガラパゴスハオリムシ)は長さ2〜3mに達する大きな動物であるが、口も肛門も消化管も無い。また、石川県能登半島九十九湾の水深約25mに棲息するOligobrachia mashikoi(図2:マシコヒゲムシ)も有鬚動物であり、その大きさは体幅1mm以下、長さ10~30cmの“髪の毛”のような動物である。それでは、このような口・肛門・消化管を持たない動物はどのような方法で生きるための食物を獲得し、エネルギーを獲得ているのだろうか?一般的には、「有鬚動物は栄養体と呼ばれる組織にイオウ酸化細菌(化学合成細菌)等を細胞内共生させ、宿主は共生するイオウ酸化細菌(化学合成細菌)が炭酸固定によりつくりだした有機物(あるいは、リソソームにより消化された共生細菌)を食物(エネルギー源)として利用している。」と考えられている。すなわち、有鬚動物にとって、細胞内共生細菌が「命の源」であり、ガラパゴスハオリムシは暗黒の世界でも生きることができる。
上記2種類の有鬚動物の大きさや棲息場所は全く異なる。しかしながら、両生物ともに硫化水素(ガラパゴスハオリムシは火山性ガスに含まれる硫化水素、マシコヒゲムシは海底土壌中の硫酸還元菌の活動により発生する硫化水素)が存在する環境に棲息し、血管中に細胞外ヘモグロビンを持っている。そして、この細胞外ヘモグロビンはヒトヘモグロビンとは全く異なり、分子量約350万や約40万の巨大な超分子複合体であり、酸素と硫化水素を同時に運搬し、宿主ミトコンドリアに酸素、細胞内共生細菌に硫化水素を供給している。従って、巨大ヘモグロビンが有鬚動物と共生細菌の生命を維持するために重要な役割を担っているといっても過言ではない。
私たちは、有鬚動物マシコヒゲムシと鹿児島湾に棲息する有鬚動物Lameribrachia satsuma(サツマハオリムシ)の巨大ヘモグロビンの“働き”を分子レベルで解明すること取り組んでいる。そのために、先ず、これら巨大ヘモグロビンの立体構造の解明に取り組んだ。その結果、2005年に京都大学大学院理学研究科三木邦夫教授グループとの共同研究によりマシコヒゲムシ巨大ヘモグロビンの立体構造を解明することに成功した(論文1,2,3)。その後、私たちは、京大グループと法政大学工学部今井清博教授グループとともに、巨大ヘモグロビンの酸素協同性に関する新しい知見を得た(論文4,5)。以下にこれまでの私たちの研究で明らかにされたマシコヒゲムシ巨大ヘモグロビンの構造機能相関について概説する。詳細については、各論文を参照いただきたい。
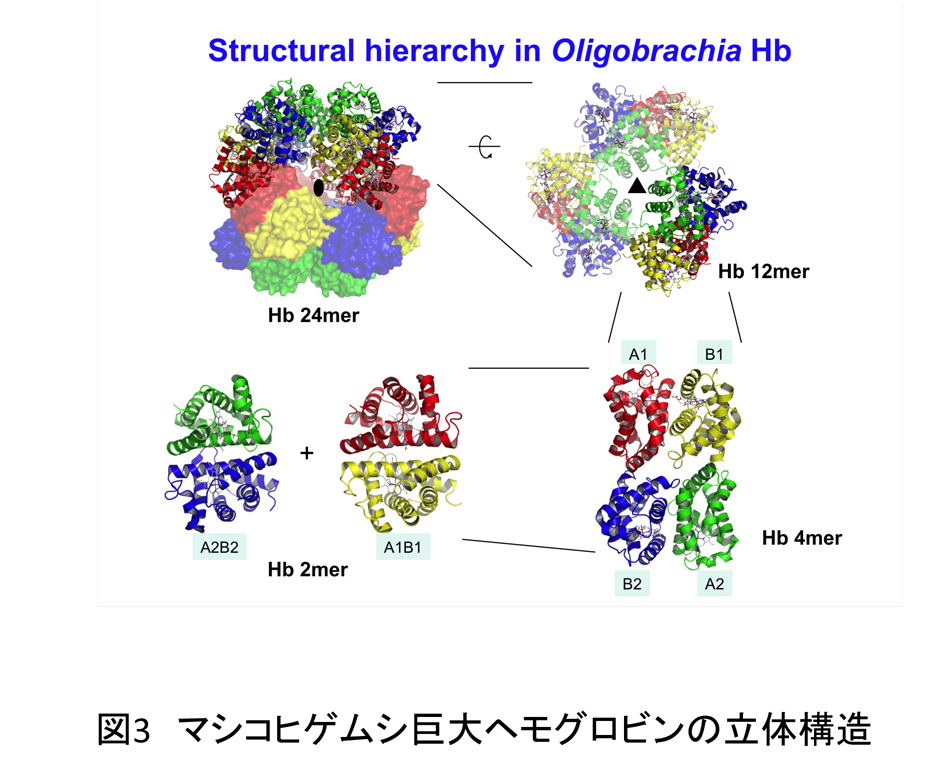
マシコヒゲムシ巨大ヘモグロビンは、4種類のグロビン鎖(A1,B1,A2,B2)で構成され、全体で24量体を形成している。全体として直径約120Åの球状構造で内部に直径約50Åの空洞が存在する。また、巨大ヘモグロビンは球を半分に分割した12量体構造が二つ会合し、24量体を形成し、12量体は4種類のグロビン鎖が各1分子集まった4量体(A1B1A2B2)が三つ会合した構造となっている。その4量体構造も、2量体構造(A1B1とA2B2)が会合した構造を持つ(図3)。 マシコヒゲムシ巨大ヘモグロビンの構造の特徴の一つは、グロビン鎖分子内およびグロビン鎖間で多くのジスルフィド結合が形成されていることである。特に、グロビン鎖間で形成されたジスルフィド結合は4量体間を結び、半球状の12量体構造の淵に沿うように配置されていて、大な複合体構造を安定的に保っている。
ヘモグロビンが酸素運搬体として機能するためには、ヘム鉄は還元型でなければならない。ヒトヘモグロビンでは、そのヘム鉄は酸化されやすいため、赤血球細胞内にはヘム鉄を還元する酵素系が機能している。それでは、マシコヒゲムシ巨大ヘモグロビンのような血管中に溶解している細胞外ヘモグロビンでは酸化されたヘム鉄はどのような方法で還元されるのだろうか?マシコヒゲムシ巨大ヘモグロビンを精製するとき、事前に血液をCOで処理しなくても、メト型ヘモグロビンは殆ど生成しない。すなわち、マシコヒゲムシ巨大ヘモグロビンのヘム鉄は酸素存在下でも殆ど酸化されないので、細胞外でも酸素運搬体として安定に機能できると思われる。現在、メト化されにくい理由を検討しているが、ヒトヘモグロビンとマシコヒゲムシ巨大ヘモグロビンでは、水分子のヘム鉄への接近しやすさが異なることが立体構造から示唆されている。次に、マシコヒゲムシ巨大ヘモグロビンの酸素協同性に関する最近の研究成果を紹介する。これまでに明らかになったことは、50mM Mgイオンや50mM Caイオンが存在すると酸素協同性が大きく上昇し、同時に酸素親和性も高くなることである(論文4)。興味あることに、マシコヒゲムシの血液中には、50mMとほぼ同じ濃度のMgイオンと Caイオンが溶解している。また、脱酸素化型*(*4種類のグロビン鎖のうち3種類のグロビン鎖のヘム鉄が酸化され、酸素が結合していない状態)と酸素化型(4種類のグロビン鎖のすべてのヘムに酸素が結合している)の立体構造を比較すると、酸素の結合によりヘムのプロピオン酸が僅かに動き、その変化を隣接するグロビン鎖のFへリックスが感知することが立体構造から示唆されている(論文5)。
以上の様に、ヒトヘモグロビンとマシコヒゲムシ巨大ヘモグロビンでは、酸素協同性の機構は大きく異なる。さらなる詳細な酸素協同性機構を明らかにするためは、金属結合型と脱酸素化型の立体構造が必須である。現在、1.6Åの高い分解能を持つ金属結合型マシコヒゲムシ巨大ヘモグロビンの立体構造の解明に成功し、その研究成果を論文投稿中である。
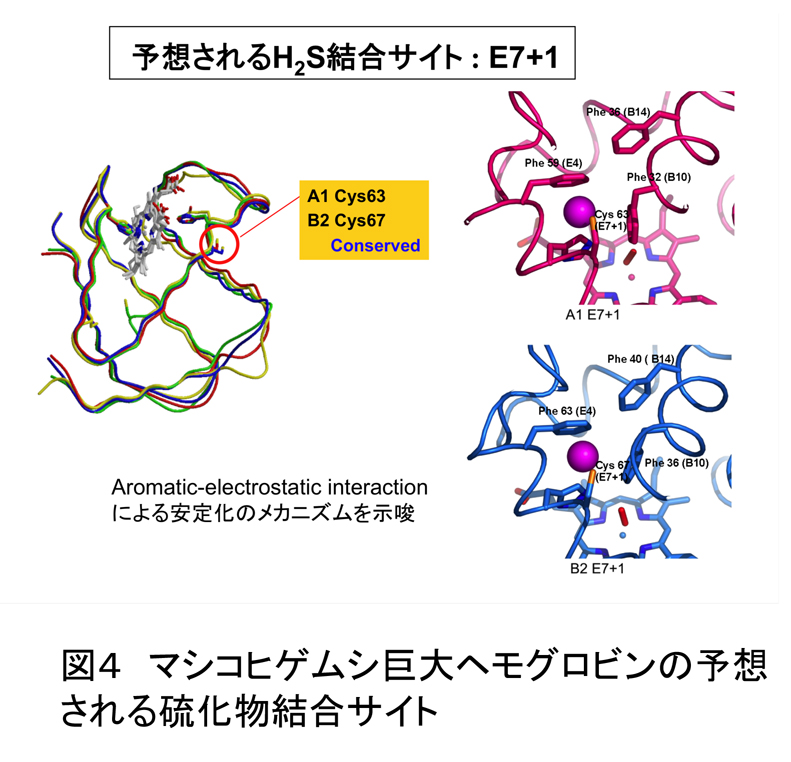
マシコヒゲムシ巨大ヘモグロビンに硫化物が結合することは、定量的な実験から明らかとなっている(論文3)。その際、ヘモグロビンの分光学的性質に変化は無い。このことは、硫化物はヘムには結合せず、蛋白質に結合していることを強く示唆する。それでは、どのアミノ酸に結合しているだろうか?これまでの研究より,硫化物を結合する機構として,フリーのシステイン残基の関与が提唱されていた。これらのシステイン残基は硫化物を結合する巨大ヘモグロビンを持つ,近縁の種間で保存されている。また、マシコヒゲムシ巨大ヘモグロビ ンの水銀化合物置換体の結晶構造中,硫化水素結合部位と提案されていたシステイン残基の全てについて,水銀化合物が結合していた。これらの実験結果は、硫化物がこのシステイン残基のイオウ原子と共有結合している可能性を強く示唆する。さらに、興味あることに、このシステイン残基の周囲はフェニルアラニンに囲まれており、硫化物結合に際して、aromatic-electrostatic interaction による安定化が推測されている(論文3)。巨大ヘモグロビンの硫化物結合部位を同定するためには、硫化物結合型の立体構造の解明がもっとも信頼できるデータだと思われるが、残念ながら、未だ、硫化物結合型ヘモグロビンの結晶の調製に成功していない。
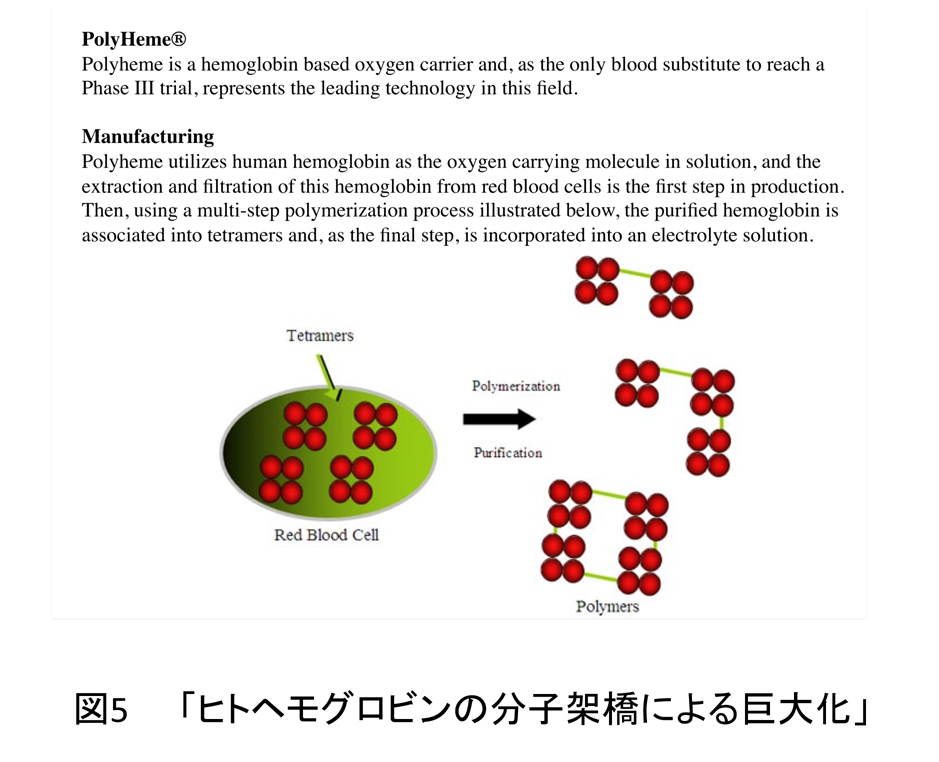
ヒトヘモグロビンは赤血球細胞内では極めて高い濃度で存在するが、もし、それらが血管中に溶解し濃度が薄くなると(具体的には約50μM以下)、α2β2の4次構造が崩壊し、ヘモグロビンとしての機能を失うことが知られている。有鬚動物巨大ヘモグロビンが血管中でヘモグロビンとして機能するためには、どんなに薄い濃度でも4量体構造を保持することが必要であり、そのために多量体化しているのかもしれない。事実、「ヒトヘモグロビンの分子架橋による巨大化」が人工血液の開発方法の一つとして注目されている(図5)。
1) Taro Nakagawa, Seiko Onoda, Masaaki Kanemori, Yuichi Sasayama and Yoshihiro Fukumori (2005) Purification, characterization and sequence analyses of the extracellular giant hemoglobin from Oligobrachia mashikoi. Zoological Science,22,283-291.
2) N.Numoto, T.Nakagawa, A.Kita, Y.Sasayama, Y.Fukumori and K.Miki (2005) Crystallization and preliminary X-ray crystallographic analysis of extracellular giant hemoglobin from pogonophoran Oligobrachia mashikoi. Biochim.Biophys.Acta, 1750, 173-176.
3) N.Numoto, T.Nakagawa, A.Kita, Y.Sasayama, Y.Fukumori and K.Miki (2005) Srructure of an extracellular giant hemoglobin of the gutless beard worm Oligobrachia mashikoi. Proc.Natl.Acad.Sci, USA, 102, 14521-14526.
4) Yayoi Aki, Taro Nakagawa, Masako Nagai, Yuichi Sasayama, Yoshihiro Fukumori, Kiyohiro Imai(2007) Oxygenation Properties of Extracellular Giant Hemoglobin from Oligobrachia mashikoi. Biochem. Biophys. Res. Comm., 360, 673-678.
5) Numoto N, Nakagawa T, Kita A, Sasayama Y, Fukumori Y, Miki K (2008) Structure of the partially unliganded met state of 400 kDa hemoglobin: Insights into ligand-induced structural changes of giant hemoglobins. Proteins, 73, 113-125.
6) 福森義宏(2004) ハオリムシ・ヒゲムシの巨大ヘモグロビンの秘密 遺伝 58巻 8-11.
7) 沼本修孝、福森義宏、三木邦夫(2006) 巨大ヘモグロビンの謎に迫る―X線結晶構造解析で構造と機能を解き明かせ! 化学, 61:30-34.
8) Nobutaka Numoto,Taro Nakagawa,Akiko Kita,Yuichi Sasayama,Yoshihiro Fukumori and Kunio Miki (2009) Structural Basis for the Heterotropic and Homotropic Interactions of Invertebrate Giant Hemoglobin, Biochemistry, 47, 11231–11238
9) 沼本修孝,中川太郎,福森義宏,三木邦夫 巨大ヘモグロビンの構造と機能 日本血液代替物学会会誌「人工血液 Artificial Blood」印刷中.